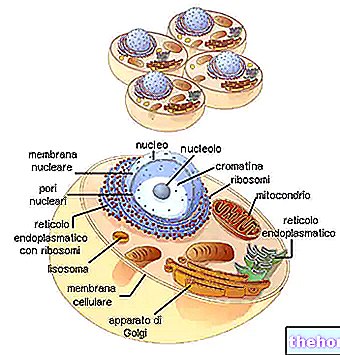
For at der er en overensstemmelse mellem informationen om polynukleotidet og polypeptidets oplysninger, er der en kode: den genetiske kode.
De generelle egenskaber ved den genetiske kode kan opføres som følger:
Den genetiske kode består af trillinger og er uden indre tegnsætning (Crick & Brenner,).
Det "blev dechiffreret ved brug af" åbne celle -oversættelsessystemer "(Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
Det er stærkt degenereret (synonymer).
Organiseringen af kodetabellen er ikke utilsigtet.
Trillinger "nonsens".
Den genetiske kode er "standard", men ikke "universel".
Når man ser på tabellen med den genetiske kode, skal det huskes, at det refererer til oversættelsen af "RNAm til polypeptid, for hvilket de involverede nukleotidbaser er A, U, G, C. Biosyntesen af en polypeptidkæde er translationen af nukleotidsekvensen i sekvens aminosyre.

Hver basistriplet af RNAm, kaldet codon, har den første base i venstre kolonne, den anden i den øverste række, den tredje i den højre kolonne. Lad os f.eks. Tage tryptophan (dvs. Try), og vi ser, at det tilsvarende kodon vil være, i rækkefølge, UGG. Faktisk inkluderer den første base, U, hele rækken af kasser øverst; i dette identificerer G boksen til højre og den fjerde linje i selve boksen, hvor vi finder skrevet Try. Tilsvarende for at syntetisere tetrapeptidet Leucine-Alanine-Arginine-Serína (symboler Leu-Ala-Arg-Ser) kan vi finde kodonerne UUA-AUC-AGA-UCA i koden.

På dette tidspunkt skal det dog bemærkes, at alle aminosyrer i vores tetrapeptid er kodet (i modsætning til tryptophan) af mere end et kodon. Det er ikke tilfældigt, at vi i det netop rapporterede eksempel har valgt de angivne kodoner.Vi kunne have kodet det samme tripeptid med en anden RNAm-sekvens, såsom CUC-GCC-CGG-UCC.
I første omgang fik det faktum, at en enkelt aminosyre svarede til mere end en trilling, en betydning af tilfældighed, også udtrykt i valget af degenerering af koden, der blev brugt til at definere fænomenet synonymi. På den anden side tyder nogle data på, at tilgængeligheden af synonymer, der kan henvises til forskellig stabilitet af den genetiske information, slet ikke er tilfældig.Dette synes også at blive bekræftet ved at finde en anden værdi af A + T / G + C -forholdet i de forskellige udviklingsstadier. For eksempel i prokaryoter, hvor behovet for variabilitet ikke opfyldes af reglerne for mendelisme og neo-mendelisme, har A + T / G + C-forholdet en tendens til at stige. Den deraf følgende lavere stabilitet i lyset af mutationer giver større muligheder for variationer tilfældigt fra genmutation.
I eukaryoter, især i multicellulære celler, hvor det er nødvendigt, at cellerne i den enkelte organisme alle holder det samme arvelige arv, har A + T / G + C -forholdet i DNA en tendens til at falde, hvilket reducerer muligheden for somatiske genmutationer .
Eksistensen af synonyme kodoner i den genetiske kode rejser, som allerede er nævnt, problemet med mangfoldigheden eller ikke af anticodons i RNAt.
Det er sikkert, at der er mindst et RNAt for hver aminosyre, men det er ikke lige sikkert, om et enkelt RNAt kan binde til et enkelt kodon eller kan genkende synonymer ligegyldigt (især når disse kun er forskellige for den tredje base).
Vi kan konkludere, at der i gennemsnit er tre synonyme kodoner for hver aminosyre, mens anticodons er mindst en og ikke mere end tre.
I erindring om, at gener er tænkt som enkelte strækninger af meget lange polynukleotidsekvenser af DNA, er det klart, at begyndelsen og slutningen af det enkelte gen nødvendigvis skal være indeholdt i hukommelsen.
BIOSYNTESE AF PROTEINER
I forskellige dele af DNA'et er der åbning af dobbeltkæden og syntesen af de forskellige typer RNA.
Under indlæsningstrinnet binder RNAt til aminosyrerne (tidligere aktiveret af ATP og af det specifikke enzym). Det biosyntetiske "maskineri" er ikke i stand til at "rette" forkert indlæste tRNA'er.
RNAr deler sig derefter i de to underenheder og giver ved binding til de ribosomale proteiner anledning til samling af ribosomerne.
RNAmet, der passerer gennem cytoplasmaet, binder sig til ribosomerne og danner polysomet.Hvert ribosom, der flyder på budbringeren, er gradvist vært for RNAt, der er komplementært til de relative kodoner, tager aminosyrerne og binder dem til polypeptidkæden i dannelse.
Det relativt stabile RNAt kommer tilbage i cirkulationen. Ribosomerne bruges også igen og frigiver det allerede samlede polypeptid.
Budbringeren, mindre stabil, fordi det hele er monokatenært, spaltes (af ribonukleasen) i de bestanddelte ribonukleotider.
Cyklussen fortsætter således og syntetiserer efter hinanden polypeptiderne på messenger -RNA'erne leveret af transkriptionen.