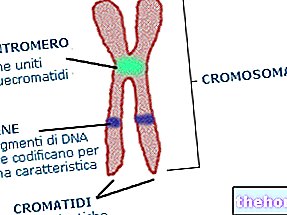
NOTER OM EMBRYOLOGI
Embryologi studerer rækkefølgen af udviklingsformer fra zygoten til organismen udstyret med alle dens organer og systemer.
I denne henseende er det værd at huske sondringen mellem udvikling (rækkefølge af strukturelle og organisatoriske faser med stigende kompleksitet) og vækst, der især er beregnet i kvantitativ forstand.
I hvirveldyr metazoans vi vidne, stigende i den evolutionære serie op til mennesket (gennem cyclostomes, fisk, padder, krybdyr, fugle og pattedyr), udseendet af voksne former for stigende kompleksitet, som komplikationen af stadierne af embryonal udvikling.
I begyndelsen er zygoten, der altid er udstyret med reservemateriale, opdelt (ved efterfølgende mitose) i 2, derefter 4, derefter 8 osv. Celler kaldet blastomerer uden vækst, indtil den når artens normale kerne / cytoplasmiske forhold .
Denne indledende segmentering kan følge forskellige mønstre afhængigt af mængden og fordelingen af deutoplasmaet.
I begyndelsen er deutoplasmaet knap ("oligolecytiske æg"), så segmenteringen er total og giver anledning til meget forskellige blastomerer. Efterhånden som embryoets kompleksitet øges, tager det mere tid og materiale, før dets udvikling tillader det at begynde. selvstændigt liv. Dette kræver en stigning i deutoplasma ("telolecytiske æg"), som har en tendens til at blive arrangeret i en del af zygoten. Dette forårsager en stigende "anisotropi", som er knyttet til ændringer af segmenteringen, styret af to generelle principper:
- Hertwigs lov siger, at ved mitose har den achromatiske spindel (hvis ækvator bestemmer dattercellernes opdelingsplan) en tendens til at blive arrangeret i retning af cytoplasmas største længde;
- Balfours lov siger, at segmenteringshastigheden er omvendt proportional med mængden af deutoplasma.
Vi ser da, at segmenteringen allerede i cyklostomer og i fisk er ulige, med en hurtigt segmenteret dyrestang (som vil give embryonets øvre strukturer) og en æggeblommestang, som vil indeholde det meste af reservematerialet. Denne tendens er endnu større . anisotrop hos padder (hvor det er nødvendigt at forberede de organer, der er ansvarlige for luftåndning), hvor æggeblommestangen, mens den langsomt segmenteres, forbliver relativt inaktiv og ender med at blive dækket af celler, der stammer fra den hurtigt segmenterende dyrepæl. succession af de vigtigste embryonale faser omfatter: zygote, blastomerer, morula (klynge af blastomerer svarende til en brombær), blastula (morula med regressive indre celler), gastrula (blastula, hvor cellerne på den ene side er blevet invagineret), hvor det primitive hulrum af organismen, med et eksternt cellelag (ektoderm, hvorfra nervesystemet først og fremmest vil stamme) og en int erno (entoderm), mellem hvilket et tredje lag (mesoderm) derefter vil placere sig. Fra disse lag eller "embryonale ark" vil derefter i en ordnet rækkefølge udlede alle væv, organer og systemer.
I endnu mere udviklede arter er stigningen i deutoplasma (eller "kalv") sådan, at den ikke engang kan segmenteres. Vi ser således, at segmenteringen hos fugle kun påvirker en tynd overfladisk skive, hvilket fører til en "discoblastula" og en række af fænomener, der garanterer dannelsen af embryoet på en anden måde end den ovenfor nævnte.
En yderligere stigning i deutoplasma ville sandsynligvis ikke have været mere effektiv, så hos pattedyr opnås udvikling og vækst op til evnen til selvstændigt liv med et andet system.Vi bemærker faktisk hos pattedyr, at deutoplasma kun bruges i de allerførste faser udviklingen; derefter etablerer embryoet metaboliske forbindelser til moderorganismen (med placentaens udseende) og bruger ikke længere deutoplasma, hvis overskydende elimineres. På dette tidspunkt vender æggene tilbage til at være oligolecitiske, og segmenteringen kan gå tilbage til at være total (og derfor ligner den i de tidlige stadier den for "amphioxus"), men efter morulaen fortsætter embryogenesen ifølge det mest udviklede fuglemønster med en "blastocyst" efterfulgt af implantation på livmodervæggen, så embryonets stofskifte sikres af moderorganismen (via moderkagen) frem for deutoplasma.
EMBRYO DIFFERENTIATION
Når segmenteringen af zygoten har bragt kerne / cytoplasmatisk forhold til artens norm, skal væksten også begynde, parallelt med udviklingen.Derfor starter metabolismen, med fremkomsten af nucleoli og proteinsyntese. Den således initierede proteinsyntese skyldes de gener, der er ansvarlige for de tidlige stadier af embryonal udvikling. Disse gener er deprimeret af stofferne, der findes i de forskellige blastomerer på dyret og kalvepolen. Til gengæld kan produkterne fra disse tidlige gener deprimere operonerne af generne, der er ansvarlige for de senere stadier. Produkterne fra denne anden serie af gener vil være i stand til at virke både i betydningen at konstruere nye embryonale strukturer og i den forstand at undertrykke de tidligere operoner og derepressere de følgende i en ordnet sekvens, der fører til konstruktionen af den nye organisme , takket være den genetiske information, der er akkumuleret. fra genomet gennem årtusinderne til stadig mere udviklede arter.
Haeckels berømte udtryk "" ontogeny opsummerer fylogeni "udtrykker faktisk det faktum, at de højere arter gentager i faser af embryonisk udvikling den succession, der allerede findes i de evolutionært tidligere arter.
De tidlige stadier af embryoet har en tendens til at være ens hos hvirveldyr, især op til gællernes udseende.
Hos arter, der skifter til luftåndning, absorberes gællerne derefter og genbruges (f.eks. Til dannelse af endokrine kirtler), men de genetiske oplysninger vedrørende dannelsen af gællerne bevares også hos mennesker. Dette er naturligvis et eksempel på embryonale strukturelle gener, der er til stede i genomet for alle hvirveldyr og skal forblive undertrykt efter at have fungeret i deres ontogenetiske øjeblik.
Fortolkningen af embryogenese i betydningen regulering af genhandling gør det muligt at forene de komplekse traditionelle erfaringer med eksperimentel embryologi.
Tvillinger
Zygoten og de første blastomerer, indtil proteinsyntesen begynder, er totipotente, det vil sige i stand til at give liv til en hel organisme. Til dette er eksperimenterne fra Spemann forbundet, der fik to embryoner fra kvælning af en amfibie -zygote. Et lignende fænomen optræder på grundlag af fænomenet identiske tvillinger hos mennesker, som netop af denne grund kaldes monozygotisk (MZ). Spemanns eksperimentelle tvillinger var halvt så store som normale, mens de hos mennesker er helt normale. Dette forklares, fordi de to embryoner i amfibierne måtte dele den eneste æggeblomme, der allerede var modtaget, mens embryonerne ved manden kunne modtage alt, hvad der er nødvendigt for deres udvikling og vækst.
Det skal huskes, at hos "mennesker har to tredjedele af tilfældene med tvillinger" en anden oprindelse: de stammer fra lejlighedsvis samtidig modning af to follikler, med frigivelse af to ægløsninger, som, når de befrugtes, giver to zygoter; faktisk i i dette tilfælde taler vi om dizygotiske tvillinger (DZ).
Da MZ -tvillingerne, divideret med mitose fra den enkelte zygote, har det samme genom, skal forskellene mellem dem være af miljømæssig oprindelse.I stedet er genomet for to DZ -tvillinger kun lige så meget som for to brødre. Tvillingemetode, meget udbredt inden for menneskelig genetik og også inden for sport.
I "mennesket, hvor visse etiske grunde ville forbyde eksperimentering, kan det konstateres, hvor meget enhver karakter reguleres af arvelige faktorer: Faktisk er strengt nedarvede tegn (f.eks. Blodgrupper) altid kun i overensstemmelse med MZ -tvillinger; at overensstemmelse af et træk i MZ er tæt på det for DZ, udledes det, at miljøfaktorer hersker frem for arvelige faktorer ved bestemmelsen af det fænotypiske træk.