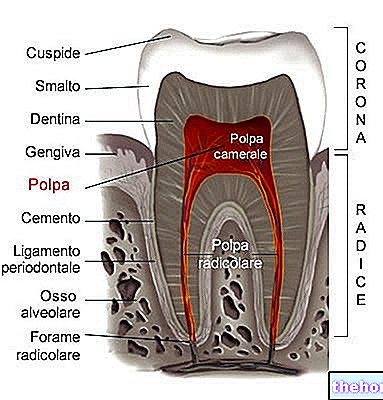
Glykogen er et makromolekyle (molekylmasse på ca. 400 millioner dalton) af α-glucose, hvor der hovedsageligt er α-1,4 glykosidbindinger og forgreninger i et forhold på 1:10 på grund af α-1,6 glykosidbindinger.
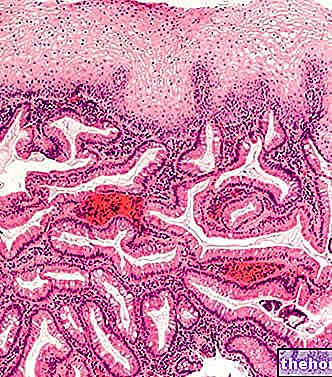
Glykogen udgør reservemateriale og nedbrydes og rekonstitueres løbende; i hele kroppens cellemasse er der omkring 100 g glykogen: det meste er i leveren, hvor det er mobilt og kan derfor bruges som reserve for andre organer (glykogen i muskler er ikke mobil).
Enzymerne, der katalyserer nedbrydning og syntese af glykogen er alle i cytoplasmaet, derfor er der brug for et reguleringssystem, der gør den ene vej inaktiv, når den anden er aktiv: hvis der er glukose tilgængelig, omdannes sidstnævnte til glykogen (anabolisme), som er en reserve, omvendt, hvis c "er påkrævet for glucose, nedbrydes glykogenet (katabolisme).

Enzymet hovedsageligt involveret i nedbrydningen af glykogen er glykogenphosphorylase; dette enzym er i stand til at spalte en glycosidisk α-1,4-binding ved hjælp af et uorganisk orthophosphat som lytisk middel: spaltningen sker på phosphorolytisk måde, og glucose 1-phosphat opnås.
Ved fem eller seks enheder fra et forgreningspunkt er glycogenphosphorylase -enzymet ikke længere i stand til at virke, derfor løsner det sig fra glykogen og erstattes af et deramificerende enzym, som er et transferase: i dette enzyms katalytiske sted c "er et" histidin, som tillader overførsel af tre saccharidenheder til den nærmeste glycosidiske kæde (histidin angriber det første carbon i et glukosemolekyle). Det netop nævnte enzym er glycosyltransferase; ved afslutningen af dette enzyms virkning er der kun én glukosenhed tilbage på sidekæden med det første kulstof bundet til det sjette kulstof i en glukose i hovedkæden Den sidste glukosenhed i sidekæden frigives ved virkningen af "enzym a-1,6 glycosidase (dette enzym udgør den anden del af det deramificerende enzym); givet at grenene i glykogen er i et forhold på 1:10 får vi ved fuldstændig nedbrydning af makromolekylet ca. 90% glucose 1-phosphat og ca. 10% af glukose.
Virkningen af de førnævnte enzymer tillader fjernelse af en sidekæde fra glykogenmolekylet; aktiviteten af disse enzymer kan gentages, indtil den fuldstændige nedbrydning af kæden sker.
Lad os overveje en hepatocyt; glukose (assimileres gennem kosten), når den kommer ind i cellen, omdannes til glucose 6-fosfat og aktiveres dermed. Glucose 6-fosfat, ved virkningen af phosphoglucomutase, omdannes til glucose 1-phosphat: sidstnævnte er en ikke-umiddelbar forløber for biosyntese; i biosyntese anvendes en aktiveret form for sukker, der er repræsenteret af sukker forbundet med et diphosphat: normalt uridyldiphosphat (UDP). Glukose 1- phosphat er derefter omdannet til UDP-glucose, denne metabolit under virkning af glykogensyntase som er i stand til at binde UDP-glucose til en ikke-reducerende ende af det voksende glykogen: forlænget glycogen fra en glucosidenhed og UDP opnås UDP omdannes af det nukleosiderede diphosphokinase-enzym til UTP, som vender tilbage til cirkulationen.
Nedbrydningen af glykogen sker ved virkningen af glykogenphosphorylase som frigiver et molekyle glukose og omdanner det til glucose 1-phosphat. Efterfølgende omdanner phosphoglucomutase glucose 1-phosphat til glucose 6-phosphat.
Glykogen syntetiseres frem for alt i leveren og musklerne: i organismen er der 1-1,2 hektar glykogen fordelt i muskelmassen.
Glykogen af en myocyt repræsenterer kun en energireserve for denne celle, mens glykogen indeholdt i leveren også er en reserve for andre væv, det vil sige, at det kan sendes som glukose til andre celler.
Glucose 6-fosfatet opnået i musklerne fra nedbrydning af glykogen sendes derefter i tilfælde af energibehov til glykolyse; i leveren omdannes glucose 6-fosfat til glukose ved hjælp af glucose 6-phosphatphosphatase (karakteristisk enzym for hepatocytter) og transporteres ind i blodbanen.
Glycogensyntase og glykogenphosphorylase virker begge på de ikke-reducerende enheder af glykogen, så der skal være et hormonalt signal, der kommanderer aktivering af den ene vej og blokering af den anden (eller omvendt).
I laboratoriet var det muligt at forlænge glykogenkæden ved at udnytte glykogenphosphorylase og bruge glucose 1-phosphat i en meget høj koncentration.
I celler katalyserer glykogenphosphorylase kun nedbrydningsreaktionen, fordi metabolitternes koncentrationer er sådan, at de skifter ligevægten i den følgende reaktion til højre (dvs. mod nedbrydning af glykogen):

Lad os se virkningsmekanismen for glykogenphosphorylasen: acetal oxygenet (som fungerer som en bro mellem glukosenhederne) binder sig til phosphorylets brint: et reaktionsmellemprodukt dannes givet af en carbokation (på glukosen, der er alt " ekstremiteter), som phosphoryl (Pi) binder meget hurtigt.
Glykogenphosphorylase kræver en cofaktor, som er pyridoxalphosphat (dette molekyle er også en cofaktor for transaminaser): den har en delvist protoneret phosphoryl (pyridoxalphosphat er omgivet af et hydrofobt miljø, der begrunder tilstedeværelsen af protoner bundet til det). Phosphorylen (Pi) er i stand til at overføre en proton til glycogenet, fordi denne phosphoryl derefter genvinder protonen fra den delvist protonerede phosphoryl i pyridoxalphosphatet. Sandsynligheden for, at phosphorylen ved fysiologisk pH mister sin proton og forbliver fuldstændigt deprotoneret, er meget lav.
Lad os nu se, hvordan phosphoglucomutase virker.Dette enzym præsenterer på det katalytiske sted en rest af phosphoryleret serin; serin giver phosphoryl til glucose 1-phosphat (i position seks): glukose 1,6-bisphosphat dannes i kort tid, derefter rephosphoryleres serin og tager phosphorylen i position et. Phosphogluco mutase kan virke i begge retninger, dvs. omdanne glucose 1-phosphat til glucose 6-phosphat eller omvendt; hvis glukose 6-fosfat produceres, kan det sendes direkte til glykolyse, i musklerne eller omdannes til glukose i leveren.
Enzymet uridylphosphogluco transferase (eller UDP glucose pyrophosphorylase) katalyserer glucose 1-phosphatoverføringsreaktionen til UTP ved vedhæftning til phosphoryl a.
Det netop beskrevne enzym er en pyrophosphorylase: dette navn skyldes, at den modsatte reaktion til den netop beskrevne er en pyrophosphorylering.
UDP -glucose, opnået som beskrevet, er i stand til at forlænge glykogenkæden med en monosaccharidenhed.
Det er muligt at få reaktionen til at udvikle sig mod dannelsen af UDP -glucose ved at fjerne et produkt, der er pyrophosphat; enzymet pyrophosphatase omdanner pyrophosphat til to molekyler orthophosphat (hydrolyse af et anhydrid) og holder på den måde koncentrationen af pyrophosphat så lav, at dannelsen af UDP -glucose termodynamisk favoriseres.
Som nævnt er UDP -glukose takket være glykogensyntases virkning i stand til at forlænge glykogenkæden.
Forvirkningerne (i et forhold på 1:10) skyldes, at når et glykogenkæde består af 20-25 enheder, griber et forgreningsenzym (der har et "histidin på sit katalytiske sted) i stand til at overføre en række 7-8 glykosidiske enheder længere nede af 5-6 enheder: således genereres en ny forgrening.
Af årsager til nervøs oprindelse eller hvis der er brug for energi på grund af fysisk anstrengelse, udskilles adrenalin fra binyrerne.
Målcellerne for adrenalin (og noradrenalin) er lever, muskler og fedtvæv (i sidstnævnte er der nedbrydning af triglycerider og cirkulation af fedtsyrer: følgelig produceres glucose i mitokondrierne 6 -phosphat, der skal sendes til glykolyse, mens glukose 6-phosphat i adipocytter omdannes til glukose ved virkningen af enzymet glucose 6-phosphatphosphatase og eksporteres til vævene).
Lad os nu se adrenalins virkemåder. Adrenalin binder til en receptor placeret på cellemembranen (af myocytter og hepatocytter), og dette bestemmer translationen af signalet udefra til inde i cellen.Proteinkinasen aktiveres, som virker samtidigt på de systemer, der regulerer syntese og nedbrydning af glykogen:
Glycogensyntase findes i to former: en dephosphoryleret (aktiv) form og en phosphoryleret (inaktiv) form; proteinkinase phosphorylerer glykogensyntase og blokerer dets virkning.
Glykogenphosphorylase kan eksistere i to former: en aktiv form, hvor en phosphoryleret serin er til stede, og en inaktiv form, hvor serin dephosphoryleres. Glykogenphosphorylase kan aktiveres af enzymet glykogenphosphorylase kinase. Glykogenphosphorylase kinase er aktiv, hvis den phosphoryleres og inaktiv, hvis den dephosphoryleres; proteinkinasen har som substrat glykogenphosphorylasekinasen, det vil sige, at den er i stand til at phosphorylere (og derfor aktivere) sidstnævnte, som igen aktiverer glykogenphosphorylasen.
Når adrenalinsignalet er overstået, skal den effekt, det har på cellen, også ophøre: phosphataseenzymer griber derefter ind på proteinarten.