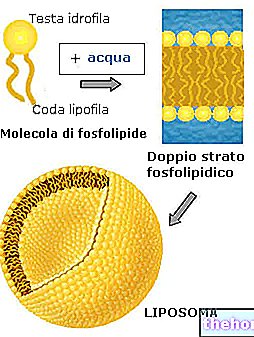
For at tale om de tyve aminosyrer, der udgør proteinstrukturerne og de modificerede, ville det være nødvendigt at beskrive mindst tolv specialiserede metaboliske veje.
Men hvorfor bruger celler så mange metaboliske veje, der kræver energi (f.eks. Til at regenerere enzymets katalytiske steder), hver med et enzymatisk arv, til at nedbryde aminosyrer? Fra næsten alle aminosyrer er det muligt via specialiserede veje at opnå metabolitter, som i mindre grad bruges til at producere energi (f.eks. Gennem glukoneogenese og ketonlegemers vej), men som frem for alt fører til dannelse af kompleks molekyler, med et stort antal carbonatomer (f.eks. fra phenylalanin og tyrosin, produceres hormoner i binyrerne, som er specialiserede til dette formål); hvis det på den ene side ville være enkelt at producere energi fra aminosyrer, på den anden side ville det være kompliceret at bygge komplekse molekyler startende fra små molekyler: aminosyrernes katabolisme gør det muligt for dem at udnytte deres skelet til at opnå større arter.
To eller tre hektogram aminosyrer nedbrydes dagligt af et sundt individ: 60-100 g af dem stammer fra de proteiner, der indføres i kosten, men over 2 hektar opnås fra den normale omsætning af proteiner, der er en integreret del af organismen ( aminosyrer af disse proteiner, der er beskadiget af redoxprocesser, erstattes af andre og kataboliseres).
Aminosyrer giver et energibidrag med hensyn til ATP: efter fjernelse af a-aminogruppen kan det resterende kulstofholdige skelet af aminosyrer efter passende transformationer komme ind i krebs-cyklussen. Når tilførslen af næringsstoffer mangler, og mængden af glukose falder, aktiveres glukoneogenese: glukoneogenetiske aminosyrer siges at være dem, der efter passende ændringer kan indføres i glukoneogenese; glukoneogenetiske aminosyrer er dem, der kan omdannes til pyruvat eller i fumarat (fumarat kan omdannes til malat, der forlader mitokondrionen og i cytoplasmaet omdannes til oxaloacetat, hvorfra phosphoenolpyruvat kan opnås). eddike-acetat.
Den netop beskrevne er et meget vigtigt aspekt, fordi aminosyrer kan afhjælpe en sukkermangel i tilfælde af umiddelbar faste; hvis fasten vedvarer, lipidmetabolismen griber ind efter to dage (fordi proteinstrukturerne ikke kan angribes for meget), er det i denne fase, at da glukoneogenese er meget begrænset, omdannes fedtsyrerne til acetylcoenzym A og ketonlegemer. Fra yderligere faste tilpasser hjernen sig også til at bruge ketonlegemerne.
Overførslen af a-aminogruppen fra aminosyrer sker gennem en transamineringsreaktion; de enzymer, der katalyserer denne reaktion, siger de faktisk transaminaser (eller aminotransferase). Disse enzymer bruger en enzymatisk kofaktor kaldet pyridoxalphosphat, som griber ind i aldehydgruppen. Pyridoxalphosphat er produktet af phosphoryleringen af pyridoxin, som er et vitamin (B6), der hovedsageligt findes i grøntsager.
Transaminaser har følgende egenskaber:
Høj specificitet for et ketoglutarat-glutamat-a-par;
De er opkaldt efter det andet par.
Transaminaseenzymer involverer altid a-ketoglutarat-glutamatparret og skelnes efter det andet involverede par.
Eksempler:
L "aspartat transaminase dvs. GOT (Glutamat-Ossalacetat Transaminase): enzymet overfører a-aminogruppen fra aspartat til a-ketoglutarat, hvorved der opnås oxaloacetat og glutamat.
L "alanintransaminase dvs. GTP (Glutamat-Pyruvat Transaminase): enzymet overfører a-aminogruppen fra "alanin til" α-ketoglutarat, hvorved der opnås pyruvat og glutamat.
De forskellige transaminaser anvender a-ketoglurat som acceptor af aminosyrenes aminogruppe og omdanner det til glutamat; mens de dannede aminosyrer bruges i ketonlegemernes vej.
Denne type reaktion kan ske i begge retninger, da de bryder og danner bindinger med det samme energiindhold.
Transaminaserne er både i cytoplasma og i mitokondrien (de er for det meste aktive i cytoplasmaet) og adskiller sig i deres isoelektriske punkt.
Transaminaserne er også i stand til at decarboxylere aminosyrer.
Der skal være en måde at konvertere glutamat tilbage til α-ketoglutarat: dette gøres ved deamination.
Der glutamat dehydrogenase det er et enzym, der er i stand til at omdanne glutamat til a-ketoglutarat og derfor omdanne aminogrupperne af aminosyrer, der findes i form af glutamat, til ammoniak. Det, der finder sted, er en redoxproces, der passerer gennem det mellemliggende a-aminoglutarat: ammoniak og α-ketoglutarat frigives og vender tilbage til kredsløbet.
Derefter passerer bortskaffelsen af aminosyrernes aminogrupper gennem transaminaserne (som er forskellige efter substratet) og glutamatdehydrogenasen, som bestemmer dannelsen af ammoniak.
Der er to typer glutamatdehydrogenase: cytoplasmatisk og mitokondriel; cofaktoren, som også er cosubstratet for dette enzym er NAD (P) +: glutamatdehydrogenase bruger enten NAD + eller NADP + som acceptor for reducerende effekt. Den cytoplasmatiske form foretrækker, men ikke udelukkende, NADP +, mens den mitokondrielle form foretrækker NAD +. Den mitokondrielle form har til formål at bortskaffe aminogrupper: den fører til dannelse af ammoniak (som er et substrat for et specialiseret enzym i mitokondrien) og NADH (som sendes til respirationskæden). Den cytoplasmatiske form virker i den modsatte retning, det vil sige, den bruger ammoniak og α-ketoglutarat til at give glutamat (som har en biosyntetisk destination): denne reaktion er en reduktiv biosyntese, og den anvendte cofaktor er NADPH.
Glutamat dehydrogenase virker, når det er nødvendigt at bortskaffe aminosyrernes aminogrupper, f.eks. Ammoniak (via urin), eller når skelettet af aminosyrer er nødvendigt for at producere energi: Dette enzym vil derfor have negative modulatorer de systemer, der er en indikation af god energitilgængelighed (ATP, GTP og NAD (P) H) og som positive modulatorer, de systemer, der angiver et behov for energi (AMP, ADP, BNP, NAD (P) +, aminosyrer og skjoldbruskkirtelhormoner).
Aminosyrer (hovedsageligt leucin) er positive modulatorer af glutamatdehydrogenase: hvis aminosyrer er til stede i cytoplasmaet, kan de bruges til proteinsyntese, eller de skal bortskaffes, fordi de ikke kan akkumuleres (dette forklarer, hvorfor aminosyrer er positive modulatorer ).
Bortskaffelse af ammoniak: urinstofcyklus
Fisk bortskaffer ammoniak ved at indføre det i vand gennem gællerne; fugle omdanner det til urinsyre (som er et kondensprodukt) og fjerner det med afføring. Lad os se, hvad der sker hos mennesker: vi har sagt, at glutamat dehydrogenase omdanner glutamat i α- ketoglutarat og ammoniak, men vi har ikke sagt, at dette kun forekommer i leverens mitokondrier.
En fundamental rolle for bortskaffelse af ammoniak gennem urinstofcyklussen spilles af mitokondrielle transaminaser.

Kuldioxid, i form af bikarbonation (HCO3-), aktiveres af biotinkofaktoren, der danner carboxybiotin, som reagerer med ammoniak for at give carbaminsyre; den næste reaktion anvender ATP til at overføre et fosfat til den carbaminsyre, der danner carbamylphosphat og ADP (omdannelsen af ATP til ADP er drivkraften til opnåelse af carboxybiotin). Denne fase katalyseres af carbamylphosphatsyntetase og forekommer i mitokondrien. Carbamylphosphat og ornithin er substrater for enzymet ornithintrans -carbamylase som konverterer dem til citrullin; denne reaktion forekommer i mitokondrier (hepatocytter). Den producerede citrullin forlader mitokondrien og går i cytoplasma under "virkningen af"argininsuccinatsyntetase: der er fusionen mellem det carbonholdige skelet af citrullin og et aspartat gennem et nukleofilt angreb og efterfølgende eliminering af vand. Argininsuccinatsyntetaseenzymet kræver et ATP -molekyle, så der er en energisk kobling: hydrolysen af ATP til AMP og pyrophosphat (sidstnævnte omdannes derefter til to molekyler orthophosphat) sker ved udvisning af et molekyle d "vand fra substratet og ikke ved virkningen af mediets vand.
Det "næste enzym er"argininsuccinase: dette enzym er i stand til at opdele argininsuccinat i arginin og fumarat i cytoplasmaet.
Urinstofcyklussen afsluttes af enzymet arginase: urinstof og ornitin opnås; urinstof bortskaffes af nyrerne (urin), mens ornitin vender tilbage til mitokondrien og genoptager cyklussen.
Urinstofcyklussen er genstand for indirekte modulering af arginin: ophobningen af arginin indikerer, at urinstofcyklussen skal fremskyndes; moduleringen af arginin er indirekte, fordi arginin positivt modulerer acetylglutamatsyntetaseenzymet. Sidstnævnte er i stand til at overføre en acetylgruppe på nitrogenet i et glutamat: N-acetylglutamat dannes, som er en direkte modulator af carbamyl-phosphosyntetaseenzymet.
Arginin akkumuleres som en metabolit i urinstofcyklussen, hvis produktionen af carbamylphosphat ikke er tilstrækkelig til at bortskaffe ornithinen.
Urea produceres kun i leveren, men der er andre steder, hvor de første reaktioner finder sted.
Hjernen og musklerne bruger særlige strategier til at eliminere aminogrupper. Hjernen bruger en meget effektiv metode, hvor et enzym bruges glutaminsyntetase og et enzym glutamase: den første findes i neuroner, mens den anden findes i leveren. Denne mekanisme er meget effektiv af to grunde:
To aminogrupper transporteres fra hjernen til leveren i et enkelt køretøj;
Glutamin er meget mindre giftigt end glutamat (glutamat udfører også neuronal overførsel og må ikke overstige den fysiologiske koncentration).
Hos fisk bringer en lignende mekanisme aminogruppen aminosyrer til gællerne.
Fra musklen (skelet og hjerte) når aminogrupperne til leveren gennem glucose-alanin-cyklussen; det involverede enzym er glutamin-pyruvat-transaminase: det tillader transponering af aminogrupper (som er i form af glutamat), omdanner pyruvat til alanin og samtidig glutamat til a-ketoglutarat i musklen og katalyserer omvendt proces i leveren.
Transaminaser med forskellige opgaver eller positioner har også strukturelle forskelle og kan bestemmes ved elektroforese (de har forskellige isoelektriske punkter).
Tilstedeværelsen af transaminaser i blodet kan være et symptom på lever- eller hjerteskade (dvs. vævsskade på lever eller hjerteceller); transaminaserne er i meget høje koncentrationer både i leveren og i hjertet: gennem elektroforese er det muligt at fastslå, om skaden er sket i leveren eller hjertecellerne.