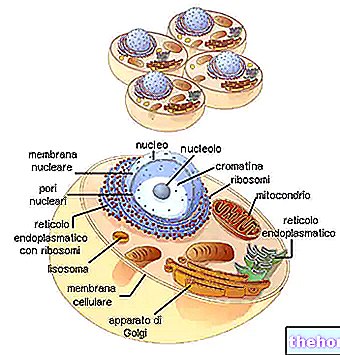
RNAm
"RNAm, eller" messenger ", tager dette navn, fordi det er ansvarligt for at bære" beskeden "af genetisk information fra det sted, hvor det er hugget (det nukleare DNA) til det sted, hvor det vil blive læst (proteinets sted syntese i cytoplasma).
Hvordan sker alt dette?
Vi har allerede set, at "aktiviteten af nukleare DNA adskiller sig i et" autosyntetisk "øjeblik (reduktion i fase S) og et" allosyntetisk "øjeblik (transskription, G1 og G2).
I begge tilfælde er vi vidne til udfoldelsen af DNA-dobbeltspiralen og "åbningen af" lynet ". Dog kan vi skelne mellem reduplikation og transkription, husk at" reduplikationsenzymet (DNA-polymerase) "løber gennem begge kæder i det øjeblik, hvor hydrogenbindinger åbnes mellem komplementære baser, mens transkriptionsenzymet (RNA-polymerase) kun går igennem en.
I erindring om, at de to DNA-kæder er "antiparallelle", og derfor på siden af åbningen begynder den ene med carbon 5 og den anden med carbon 3 af pentosen, er det nok at forestille sig, at RNA-polymerasen kun kan begynde at læse med carbon 5 for at forklare det faktum, at kun én DNA -kæde fungerer som et gen, det vil sige som en skabelon for RNA.
DNA -SEKVENS KOPIERER EN RNAm -MOLEKYLE.
det er tydeligt, at hvis kopien fandt sted på begge DNA -kæder, ville hver produceret messenger svare til en komplementær messenger med en helt anden sekvens. Hver gang cellen skulle bruge et bestemt gen, ender den med to produkter, hvoraf den ene ikke kun kan være ubrugelig, men også skadelig.
Under transkription "kopierer" RNA -polymerasen informationen i et gen til DNA'et til et mRNA -molekyle. Denne proces ligner prokaryoter og eukaryoter. En bemærkelsesværdig forskel er imidlertid, at "RNA -polymerase af eukaryoter er associeret med mRNA -kontrol af enzymer under transkription, så modifikationen foregår hurtigt efter transkriptionen begynder. Det umodificerede eller delvist modificerede produkt kaldes pre-mRNA, som når det modificeres kaldes modent RNA. [http://it.wikipedia.org/wiki/RNA_messaggero]
Transkriptionen, det vil sige udskrivning af "RNAm med DNA'et, involverer følgende fænomener: 1) afvikling af DNA-spiralen 2) åbning af" flash "; 3) tilstedeværelse af RNA-polymerase; 4) tilgængelighed af ribonukleotider af de fire typer; 5) tilgængelighed af energi til at "aktivere" og binde ribonukleotider sammen.
RNAm -molekylet syntetiseres gradvist i en sekvens bestemt ved komplementaritet med DNA'ets. For hver adenin, guanin, thymin eller cytosin i DNA'et vil blive arrangeret i henholdsvis den komplementære RNA -kæde uracil, cytosin, adenin og guanin, altid i henhold til dobbelt- og tredobbelt bindingsprincippet. Derefter løsnes RNAm -molekylet og frigives og migrerer mod cytoplasmaet, hvor det vil binde til ribosomerne for at give anledning til proteinsyntese.
RNAm-molekyler menes generelt at være enkeltkædede. Dette bekræftes af manglen på definerede relationer mellem baseparene og svarer til behovet for begrænset stabilitet.
Faktisk, hvis RNAm -molekylet var meget stabilt, ville det fortsætte med at producere det tilsvarende polypeptid på ubestemt tid, selv når det blev overdreven. På den anden side kan RNAm, der er monokatenær, let bryde ned i komponent ribonukleotider (genanvendelige), mens enhver forlænget produktion af det relative polypeptid vil blive sikret ved den fortsatte transkription af nyt RNAm.
Det skal bemærkes, at transkriptionen vedrører overførsel af information fra et 4-bogstavet alfabet til et andet 4-bogstavet alfabet (med den eneste forskel på U i stedet for T), og at den relative proces stadig forekommer for enkelte nukleotider, mens det vil være i oversættelsen, at overgangen til et alfabet på 21 bogstaver og aflæsning af nukleotiderne vil finde sted, ikke individuelt, men 3 ad gangen (i trillinger).
RNAr
RNAr, eller ribosomalt, er byggesten for ribosomer.
RNAr udskrives fra DNA, og netop fra den kanal af visse kromosomer kaldet nucleolar -organisatoren. Dette svarer til, at nucleolus er hovedlageret af RNAr, som binder til de tilsvarende proteiner. Generne, der er ansvarlige for syntesen af " RNAr "RNAr udgør en lang strækning af RNA, alligevel gentaget hundredvis af gange (dette fænomen får navnet redundans: det svarer til behovet for at intensivere produktionen af en bestemt type RNA og garantere dets produktion). Hvert gen udskriver den en kæde af ANN'er, som i tilfælde af "RNAt og RNAm.
RNAt
RNAt (transfer RNA eller transport) kaldes så, fordi det transporterer aminosyrerne (spredt i cytoplasmaet) til proteinsyntesens sted, det vil sige til det punkt, hvor ribosomet (der flyder langs "RNAm)" syr "aminosyren syrer sammen i den ordnede serie af polypeptidet. Det kaldes også RNA'er (opløseligt), fordi det er et relativt lille molekyle, der frit kan cirkulere i opløsning.
Når messenger -RNA'et via et kodon specificerer indsættelse af en bestemt aminosyre, tages dette ikke direkte fra cytoplasmaet, men aktiveres først i nærvær af et specielt enzym og ATP (som leverer energi ved at overføre det til aminosyren syre), hvorefter den binder sig til et specifikt RNAt, som bærer de reaktive steder både for at binde til aminosyren (specifikt genkende dens sidekæde), og for at fiksere sig på ribosomet og på messenger -RNA'et. RNAt, der bærer aminosyren, reagerer med messenger, da den har et særligt sted, en trilling kaldet anticodon, som er komplementær til codonet i henhold til de sædvanlige to regler for komplementaritet af nukleinsyrer.
Nukleotidsekvensen for noget RNAt er allerede blevet konstateret, hvilket generelt ser ud til at være inden for 100 -nukleotidområdet.
Alle RNAt slutter med en fast triplet kaldet CCA, som er bestemt til at binde til aminosyrens carboxylfunktion. Der er blevet fremsat forskellige hypoteser om RNAt's rumlige konformation, herunder en hårnål og en kløver. især suggestiv, fordi den har fire reaktive steder: CCA -terminalen, der binder til carboxyl (og fælles for alle aminosyrer), en "anden konstant triplet, der binder til ribosomet (også konstant), en specifik triplet, der binder til den kædespecifikke side af aminosyren og anticodon, som binder til det tilsvarende specifikke codon.






















-cause-sintomi-e-terapia.jpg)
