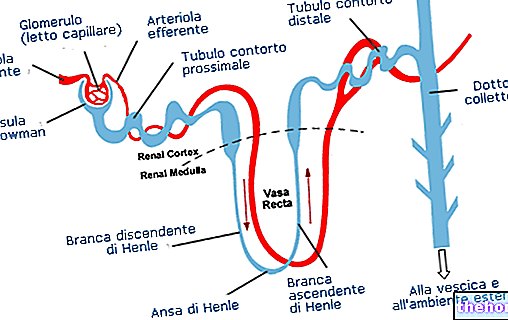
Den respiratoriske kvotient er en meget nyttig parameter til evaluering af den metaboliske blanding, der bruges i hvile eller under fysisk træning. På grund af de kemiske forskelle, der kendetegner dem, kræver fuldstændig metabolisering af fedtstoffer, proteiner og kulhydrater forskellige mængder ilt. Følgelig vil typen af oxideret energisubstrat også påvirke mængden af produceret kuldioxid.

QR = CO2 produceret / O2 forbrugt
I betragtning af at hvert makronæringsstof har en specifik QR, er det ved at evaluere denne parameter muligt at spore blandingen af næringsstoffer, der metaboliseres i hvile eller under en bestemt arbejdsaktivitet.
Respiratorisk kvote af kulhydrater
Den generiske molekylformel for et kulhydrat er Cn (H2O) n. Det følger heraf, at inden for et kulhydratmolekyle er forholdet mellem antallet af hydrogenatomer og ilt fastsat og lig med 2: 1. For at oxidere en generisk hexose (kulhydrat med seks carbonatomer såsom glucose) vil det derfor være nødvendigt seks ilt molekyler, hvilket resulterer i dannelsen af 6 kuldioxidmolekyler (C6H1206 + 602 → 6H20 + 6C02).
Den respiratoriske kvote af kulhydrater vil derfor være lig med: 6CO2 / 6O2 = 1,00
Åndedrætsværdi for lipider
Lipider skelnes fra kulhydrater ved det lavere iltindhold i forhold til antallet af hydrogenatomer. Derfor kræver deres oxidation en større mængde ilt.
Ved at tage palmitinsyre som et eksempel opdager vi, at der under oxidationen dannes 16 molekyler kuldioxid og vand til 23 forbrugte iltmolekyler. C16H32O2 + 23 O2 → 16 CO2 + 16 H2O
Den respiratoriske kvotient vil derfor være lig med: 16 CO2 / 23 O2 = 0,696
Normalt tilskrives en respirationskvotient på 0,7 lipider, idet man husker på, at denne værdi svinger fra 0,69 til 0,73 i forhold til længden af kulstofkæden, der kendetegner fedtsyren.
Respiratorisk kvotient af proteiner
Den største forskel, der adskiller proteiner fra fedt og kulhydrater, er tilstedeværelsen af nitrogenatomer. På grund af denne kemiske forskel følger proteinmolekylerne en bestemt metabolisk vej. Leveren skal først fjerne nitrogen gennem en proces kaldet deamination. Først derefter kan den resterende del af aminosyremolekylet (kaldet ketosyren) oxideres til kuldioxid og vand.
Ligesom lipider er ketosyrer relativt iltfattige. Deres oxidation vil derfor føre til dannelse af en mængde kuldioxid, der er lavere end mængden af forbrugt ilt.
Albumin, det mest rigelige protein i plasma, oxiderer i henhold til følgende reaktion:
C72H112N2O22S + 77O2 → 63CO2 + 38 H2O + SO3 + 9 CO (NH2) 2
Den respiratoriske kvotient vil derfor være lig med: 63 CO2 / 77 O2 = 0,818
QR -værdien af proteiner fastgøres konventionelt til 0,82.
Betydning af respirationskvotienten
For at imødekomme organismens energibehov bruger vi hver især forskellige metaboliske blandinger i forhold til fysisk anstrengelse. Jo mere intens dette er, jo større procentdel af oxideret glukose. Meget af energien, der produceres i hvile, stammer fra metaboliseringen af syrer. fed. Af denne grund er det rimeligt at forvente en respirationskvotient tæt på 0,7 i hvile og højere under anstrengende træning.
At udføre aktiviteter, der spænder fra absolut hvile til let aerob træning, er respirationskvotienten omkring 0,82 ± 4%. Denne dato, opnået eksperimentelt, vidner om oxidation af organismen af en blanding bestående af 60% fedt og 40% kulhydrater (under betingelser af hvile eller moderat fysisk aktivitet er proteiners energirolle ubetydelig, derfor taler vi om respiratorisk kvotient uden protein).
Hver QR -værdi svarer til en kalorieækvivalent ilt, der repræsenterer antallet af frigivne kalorier pr. Liter O2. Takket være disse data er det muligt meget præcist at spore energiforbruget ved en arbejdsaktivitet. Lad os antage, at under moderat aerob træning er respirationskvotienten målt ved gasanalyse lig med 0,86; ved at konsultere en speciel tabel finder vi, at energiækvivalenten pr. Liter forbrugt ilt er 4,875 Kcal. På dette tidspunkt for at finde ud af energien udgifter til motion vil det være tilstrækkeligt at gange liter forbrugt ilt med 4,875.
Under intens fysisk anstrengelse ændrer situationen sig radikalt, og respirationskvotienten undergår store variationer. På grund af den massive produktion af mælkesyre aktiveres adskillige hjælpemetaboliske mekanismer, såsom buffersystemer og hyperventilation.I begge tilfælde er der en stigning i eliminering af CO2, uafhængigt af oxidationen af energisubstraterne. Til tælleren (CO2 ) og holde nævneren konstant (O2) undergår respirationskvotienten en stigning, der når værdier højere end en.
Under genopretningen efter en intens aktivitet, når en del af kuldioxid bruges til at reformere bikarbonatreserverne, falder respirationskvotienten under grænseværdien 0,70.
Det er derfor klart, at respirationskvotienten i sådanne situationer ikke ligefrem afspejler, hvad der sker på celleniveau under oxidation af energisubstrater. I disse tilfælde foretrækker respirationsfysiologerne at tale om den eksterne respirationskvotient eller forholdet mellem respiratoriske udvekslinger (R).