Levende væseners cykliske kontinuitet finder forbindelserne mellem successive generationer i reproduktionens fænomener.
Reproduktion udføres på forskellige niveauer af den evolutionære skala, i de forskellige forgreninger af grøntsags- og dyreriget, i de forskellige levende arter, med en sådan række mekanismer, der alene berettiger en hel afhandling.
En første klassificering af reproduktionsfænomenerne skal skelne encellede fra flercellede organismer, da kun i førstnævnte falder celledeling sammen med reproduktion.
I flercellede kan reproduktion være agamisk eller seksuel (eller gamic).
Agamisk reproduktion, relativt mindre hyppig, er baseret på mekanismen for mitose, så artens variabilitet snarere er betroet til gentagen forekomst af mutationer.
Der er også forskellige mekanismer, såsom strobilisering, sporulation osv., Mens vi i planter finder regenerative former velkendte i landbruget (stiklinger, lagdeling osv.).
Den mest udbredte reproduktionsmekanisme i de højere former er imidlertid den seksuelle, svarende til forekomsten af meiose, dannelsen af kønsceller og deres fusion i zygoten (befrugtning).
Hos primitive arter er kønscellerne ikke morfologisk differentierede: i dette tilfælde taler vi om isogametia. Der er imidlertid altid to serier af kønsceller, der er identificeret med symbolerne (+) og (-), og befrugtning kan kun finde sted fra "mødet mellem kønsceller med modsat tegn: der er derfor en biologisk forskel, der endnu ikke er udtrykt morfologisk.
Efterhånden som den evolutionære skala skrider frem, fremkommer morfologisk og funktionel differentiering, hvor en type kvindelige kønsceller generelt er udstyret med rigeligt reservemateriale (deutoplasma eller kalv, som vil sikre udviklingen af embryoet, indtil det kan være metabolisk uafhængigt) og en type mandlige kønsceller udstyret med motilitet til at nå kvindelige. Gameter er altid haploide og er et resultat af meiose. Deres fusion giver anledning til diploid zygote.
Mellem meíosis og zygote kan der forløbe en række haploide cellegenerationer, ligesom der mellem zygote og meiose kan forløbe en række diploide cellegenerationer, hvor de forskellige sorter af generative cyklusser udtrykkes ved skiftende generationer.
En diplonte -art (med en diploid organisme) er kendetegnet ved gametisk meiose: meiose producerer direkte kønscellerne, som ved fusion straks rekonstituerer den diploide tilstand. Dette er det udbredte tilfælde af Metazoans, herunder mennesket.
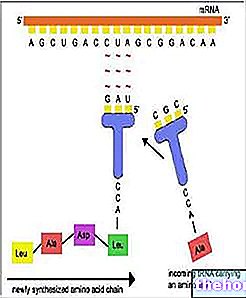
Gametogenese
Under hensyntagen til reproduktionen med gametisk meiose, som den forekommer hos mennesker, lad os prøve at præcisere, hvordan meiose passer ind i gametogenese (dannelse af gameter).
I embryonisk udvikling af mandlig og kvindelig gametogenese (kaldet spermatogenese og oogenese) er der en tidlig differentiering mellem cellerne, der er bestemt til at danne kroppen (somatisk linje) og dem, der er bestemt til at producere kønsceller (kimlinje). De oprindelige celler i kimlinjen kaldes protogoner. Med differentiering af gonaden i mandlig eller kvindelig forstand er der differentiering af kimcellerne henholdsvis til spermatogonia og ovogonia.
Når man ser på spermatogenese, ser vi, at der er en række cellegenerationer i spermatogonien, som fortsætter gennem hele livet. Kun en del af den således kontinuerligt producerede spermatogoni adskiller sig fra den normale mitotiske cyklus, og den meiotiske begynder i stedet.
Kimcellen, hvori meiosen begynder (reduplikation og derefter første division) kaldes første ordens spermatocyt; dens division giver anledning til to andenordens spermatocytter, som med anden division giver anledning til i alt fire spermatider.
Vi kan gradere reduktionen af kromosomkittet fra 4n af første ordens spermatocytter (efter reduplikation er der fire kromatiler for hvert par homologer) til 2n af anden ordens spermatocytter og til n af spermatider, som allerede set ved undersøgelse meiose, som Således konkluderer vi. Spermatiderne er derfor allerede haploide, men de er endnu ikke modne kønsceller. Fra strukturen i en haploid type celle forvandler funktionel modning (kaldet spermiohistogenese) spermatiderne til spermatozoer, det vil sige modne mandlige kønsceller.
I kvindelig gametogenese (eller oogenese) er der flere forskelle. Først og fremmest er antallet af gameter, der skal forberedes, meget mindre. Det anslås, at omkring 5 X 105 ovogoner fremstilles i kønskirtlerne hos en hun af den menneskelige art; af disse er kun omkring 400 interesserede i follikelmodning og efterfølgende nedbrydning i en cyklus, der normalt kun påvirker en follikel om måneden i en frugtbar periode på cirka 35 år.
Det forskellige antal kønsceller tilberedt i de to køn svarer til forskellen i funktion og adfærd, der allerede er nævnt: Sædcellerne er små, mobile og talrige i forhold til behovet for at søge efter ægget og den lave sandsynlighed for at finde det; æggene er store, inaktive og få, i forhold til funktionen af at garantere embryoet reservematerialet og den beskyttelse, der ydes dem ved intern befrugtning (naturligvis, især ved ekstern befrugtning, skal æggene også være mere talrige).
Behovet for at forsyne kønsceller med reserve-materiale svarer til tilstedeværelsen i "oogenesen" af en fase af standsning af meiose, hvor kromosomerne delvist despiraliseres. Vi observerer derefter de såkaldte "fjerkromosomer", hvor en serie af ekstroversioner identificerer de træk, hvor de gener, der er ansvarlige for syntesen af deutoplasma, er deprimerede.
Det mindre antal, der kræves for hunlige kønsceller, svarer også til det faktum, at af de fire haploide celler, der produceres af meiose, kun én modtager hele reservematerialet og bliver til en gamette, mens de tre andre (polocytterne eller polære legemer), der kun indeholder kromosomalt materiale, kan ikke give anledning til zygoter og embryoner og er bestemt til at gå tilbage.
Befrugtning
Befrugtning, det vil sige mødet mellem den mandlige og kvindelige kønscelle, kan udføres på meget forskellige måder.I dyreriget observerer vi overgangen fra ekstern befrugtning (kønsceller udsat for enhver miljømæssig risiko og derfor nødvendigvis meget talrig i de to køn) til intern befrugtning, som forældres omsorg yderligere er knyttet til pattedyrs metaboliske forhold mellem mor og foster .
Befrugtning, når mødet mellem kønsceller af det modsatte køn har fundet sted, skal finde sted og garanterer to betingelser: specificitet og unikhed. Det vil sige, at det skal sikres, at sædcellen er af samme art som ægget, og at når den første kommer ind, kommer ingen andre ind.
Specificitet sikres af de biokemiske egenskaber ved akrosomet og æggets overflade. Faktisk tales der om reaktioner mellem «fertilisiner» og «antifertilisiner», med specificitet, der kan sammenlignes med mødet mellem enzymer og substrat.
Befrugtningens unikke karakter garanteres ved en ændring af æggets overfladestruktur («kortikal reaktion»), der begynder lige i øjeblikket af den første specifikke fertilisin / antifertilisin reaktion; efter denne reaktion ændres æggemembranen, så enhver anden sædcelle, der når den, ikke længere er i stand til at starte den specifikke befrugtningsreaktion.
Efter befrugtningen forbliver spermotozoens hale ekstern i forhold til ægget, mens det kromosomale materiale trænger ind i det. Dette, kaldet "mandlig pronucleus", slutter sig til ægets "kvindelige pronucleus" og danner dermed zygotens diploide kerne.