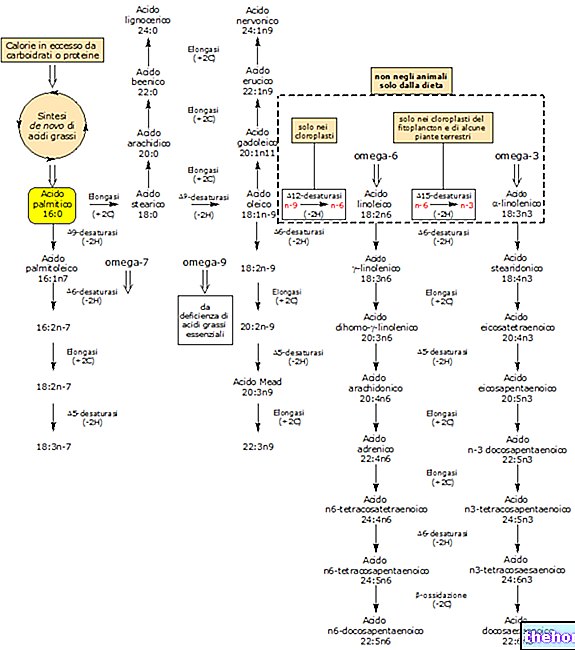
Hvis en celle arbejder under anaerobe forhold, producerer den energi ved at omdanne glukose til laktat og bortskaffer sidstnævnte gennem Cori -cyklussen; hvis der er ilt tilgængeligt (derfor under hvileforhold), forbruges mere end 90% af glukosen aerobt og kun de resterende 10%, anaerobt. Når der er behov for mere ATP, end hvad den aerobe vej er i stand til at levere (for eksempel når muskler er under stress), så leveres den ekstra forsyning af anaerob metabolisme (vi er under iltmangel: åndenød, træthed ved vejrtrækning osv.): det er nødvendigt at fremskynde dette stofskifte ved at omdanne laktat (som opnås ved glykolyse) til glucose gennem glukoneogenese.
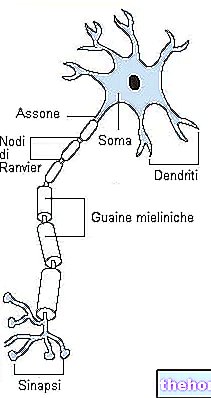
Aerob metabolisme udvikler sig i mitokondrierne.
Det første enzym, der opstår i aerob metabolisme, er pyruvat dehydrogenase; Det er mere præcist at sige, at pyruvatdehydrogenase er et enzymkompleks frem for et enzym, da det er et aggregat på 48-60 proteinenheder med tre katalytiske steder, der virker i rækkefølge.
Pyruvatdehydrogenase katalyserer følgende reaktion (redox):
Pyruvat + NAD + + CoA-SH → Acetyl CoA + NADH + H + + CO2
CoA-SH er coenzym A: det er et derivat af pantothensyre; acetylcoenzym A er en thioester. Dette er en redoxproces, fordi det første carbon af pyruvat går fra oxidation nummer tre til oxidation nummer fire (det oxiderede) og det andet carbon af pyruvat går fra oxidation nummer to til oxidation nummer tre (det oxiderede). Derefter oxideres pyruvat (det mister to elektroner totalt), og NAD reduceres.
Som nævnt har pyruvatdehydrogenase tre typer enzymatisk aktivitet, hver understøttet af sin egen katalytiske cofaktor:
- thiaminpyrophosphat (det er et derivat af vitamin B1); det er aktivt i deprotoniseret form: en carbanion dannes.
- lipoamid (det er et derivat af liponsyre); det indeholder en meget reaktiv disulfidbro.
- flavin adenin dinucleotid (det er et derivat af vitamin B2); det er et nukleotid med redoxegenskaber: dets redoxcenter består af flavinen.
I eukaryote celler forekommer aerob metabolisme i specialiserede organeller i cellen, som er mitokondrier; i bakterier forekommer metabolismen af glukose og andre arter i cellen, men der er ingen specialiserede organeller.
Når pyruvat kommer ind i en mitokondrion, udsættes det for "virkningen af pyruvatcarboxylase, hvis der er behov for at udføre glukoneogenese (for at rekonstruere udgangsmaterialet), eller det kan udsættes for pyruvatdehydrogenase, hvis det er nødvendigt at producere energi: "acetylcoenzym A, der dannes ved aerob metabolisme, stimulerer virkningen af pyruvatcarboxylase, derfor fremmer det gluoconeogenese og reducerer virkningen af pyruvatdehydrogenase.
Lad os nu se, hvordan pyruvat dehydrogenase virker; først og fremmest er der en decarboxylering af pyruvat ved virkningen af thiaminpyrophosphat.
Et surt miljø kan hæmme aerob metabolisme, fordi den anioniske form af thiaminpyrophosphat er aktiv, hvilket ville blive protoneret ved surt pH, og decarboxylering ikke ville forekomme.
En decarboxylering er en vanskelig reaktion, da en carbon-carbon-binding skal brydes; i dette tilfælde er reaktionen termodynamisk begunstiget af det faktum, at reaktionsmellemproduktet (hydroxyethyl-thiaminpyrophosphat) giver resonans (molekylets p-elektroner delokaliseres): hydroxyethyl-thiamin-pyrophosphatet findes i tre mulige former (resonans) og det gør den ret stabil. Desuden overlever hydroxyethyl-thiaminpyrophosphatet i anionisk form i tilstrækkelig lang tid til at kunne interagere med lipoamidets disulfidbro (anden katalytiske cofaktor af pyruvatdehydrogenase); disulfidbroen er en oscillerende arm (den er placeret ved ende af en lang fleksibel kæde) og kan bevæge sig fra et katalytisk sted til et andet i enzymkomplekset.
Derefter binder lipoamidet gennem disulfidbroen hydroxyethyl-thiaminpyrophosphatet: acetyllipamid opnås.Dette er den første fase af en transacetyleringsreaktion katalyseret af det første enzym i pyruvatdehydrogenasekomplekset; i denne fase blev det brudt en binding mellem hydroxylgruppen og thiaminpyrophosphat, der vendte tilbage til sin oprindelige form: en redoxreaktion fandt sted, hvor disulfidbroen virkede som en oxidant (de to svovlatomer reduceret) mod hydroxylgruppen, som den oxiderede til acetyl.
Efter denne fase bevæger lipoamidets oscillerende arm sig og nærmer sig det andet enzym af pyruvatdehydrogenase, som udfører den sande transacetylaseaktivitet ved at bære acetylgruppen med sig: den anden fase af transacetyleringsreaktionen katalyseret af det andet enzym finder sted; på denne måde har vi opnået acetylcoenzym A. Det er nu nødvendigt at genoprette lipoamidet, der er i reduceret form: det tredje enzym af pyruvatdehydrogenase griber ind, som redoxerer lipoamidet og overfører dets elektroder til FAD, som reduceres til FADH2. Parret FAD / FADH2 kan fungere som et redox -par i to forskellige monoelektroniske stadier eller i et enkelt bielektronisk trin.
FADH2 giver straks sine elektroner til NAD + for at opnå FAD og NADH + H +.
Acetylcoenzym A, opnået som beskrevet, er udgangsproduktet for Krebs -cyklussen (eller cyklussen af tricarboxylsyrer).















.jpg)