Syntesen af fedtsyrer starter fra acetylcoenzym A og svarer nogenlunde til den omvendte vej for deres nedbrydning; ved syntese af fedtsyrer tilføjes en række bikarbonatfragmenter til udgangsacetylcoenzym A.
Syntesen af fedtsyrer er fuldstændig cytoplasmatisk (dvs. de enzymer, der katalyserer denne syntese, findes i cytoplasmaet). Acetylcoenzym A, der anvendes i cytoplasmaet til syntese af fedtsyrer, er af mitokondriel oprindelse: en lille del transporteres gennem carnitin ved hjælp af to acyltransferaseenzymer (et cytoplasmatisk og et mitokondrielt) og et translocaseenzym. En del af acetyl coenzym A fra mitokondriel oprindelse opnås via en specialiseret rute: citratlyase (navnet stammer fra det første enzym på denne vej).
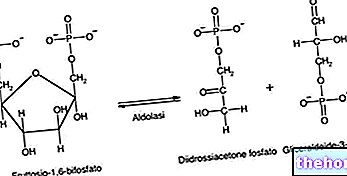
Acetylcoenzym A til stede i mitokondrier stammer fra glykolyse efter virkningen af pyruvatdehydrogenase; Acetylcoenzym A gennemgår virkningen af enzymet citratsyntase: dette enzym katalyserer dannelsen af citrat ved reaktionen af acetylcoenzym A med oxaloacetat. Hvis krebscyklussen er i stand til at opfylde energibehovet, starter det med citrat (mængden unødvendig i krebs -cyklussen) kan forlade mitokondrierne og nå cytoplasmaet, hvor citratlyaseenzymet, der bruger energi, omdanner det tilbage til acetylcoenzym A og oxaloacetat. På denne måde er det muligt at have acetylcoenzym A tilgængeligt i cytoplasmaet; oxaloacetatet, der dannes, skal dog returneres til mitokondrierne for igen at være tilgængeligt for citratsyntaseenzymet.
Oxaloacetatet omdannes derefter til malat ved enzymets virkning malat dehydrogenase cytoplasmatisk (et cytoplasmatisk NADH bruges): malatet er en permeabel metabolit og kan genindtræde i mitokondrierne, hvor det under virkningen af mitokondrielt malat dehydrogenaseenzym omdannes til oxaloacetat (der opnås også et NADH); den cytoplasmatiske patient kan alternativt undergå virkningen af det æbletiske enzym, der udfører en decarboxylering og dehydrogenering, der skal omdannes til pyruvat. Æblet -enzymet virker på NADP + (det ligner nicotinamid -adenindinukleotid, men i modsætning til dette har det en fosforsyret gruppe på den anden hydroxylgruppe på en af de to ribosenheder) derfor produceres NADPH i passagen fra malat til pyruvat ( som bruges i biosyntese) Pyruvat kommer derefter ind i mitokondrierne, hvor det omdannes til oxaloacetat ved virkningen af pyruvatcarboxylase eller til acetylcoenzym A gennem pyruvatdehydrogenase.
Lad os se et eksempel: otte molekyler acetylcoenzym A er nødvendige for at syntetisere palmitinsyre (kæde med seksten carbonatomer), men kun et af dem bruges som sådan: syv molekyler acetylcoenzym A omdannes til malonylcoenzym A af "enzymet" acetylcoenzym A carboxylase (dette enzym bruger et CO2 -molekyle og har biotin som en kofaktor).
Acetylcoenzym A -carboxylaseenzymet kan eksistere i en næsten inaktiv dispergeret form og en aktiv aggregatform (ca. tyve enheder); overgangen fra den dispergerede til aggregatformen sker, når der i cytoplasmaet er en "høj koncentration af citrat: citrat er en positiv modulator af acetylcoenzym A carboxylase -enzymet.
Acetylcoenzym A -carboxylaseenzymet har andre positive (insulin) og negative (glukagon-, adrenalin- og acylcoenzym A) -modulatorer.
Vi vil analysere syntesen af fedtsyrer i bakterien escherichia coli, hvor denne syntese sker ved virkningen af syv forskellige proteiner; i eukaryote celler svarer mekanismen til syntese af fedtsyrer til bakterier, men i eukaryoter grupperes de syv enzymer, der er ansvarlige for syntesen, i to multienzymkomplekser A og B.
I bakterier koder syv forskellige gener til:
- ACP (acylbærerprotein);
- ACP-acetyltransacetylase;
- ACP.malonyltransacetylase;
- β-keto-acyl-ACP-syntase (kondenserende enzym);
- β-keto-acyl-ACP-reduktase;
- D-p-hydroxy-acyldehydratase;
- enoil-ACP redigeret.
I eukaryoter koder to gener for:
Underenhed A.
ACP;
Kondenserende enzym
β-keto-acyl-ACP-reduktase.
Underenhed B
ACP-acetyltransacetylase;
ACP-malonyltransacetylase;
D-p-hydroxy-acyldehydratase;
enoil-ACP redigeret.
De syv proteiner af Escherichia coli er arrangeret på en sådan måde, at der er en central (ACP) og de seks andre omkring den.
To sulfhydrylgrupper er involveret i dets enzymatiske virkning: en tilhørende en cystein og en tilhørende den lange arm af en phosphopanthetein; ACP binder sig til substratet, der gennem phosphopanthethein -armen sættes i kontakt med de andre enzymer, som dermed er i stand til at udføre deres enzymatiske virkning.
Acetylcoenzym A (ved hjælp af ACP-acetyltransacylase) binder sig til ACP-enzymet (mere præcist til svovl af cystein, der danner cysteylderivatet), og coenzym A frigives; ACP-malonyltransacylase griber derefter ind, hvilket katalyserer angrebet af malonyl på phosphopanthethin (også i denne proces frigives coenzym A, der oprindeligt var bundet til malonyl).
Det næste trin involverer β-keto-acyl ACP-syntasen, som er et kondenserende enzym: det tillader fusion mellem de to skeletter; malonyl decarboxyleres let, og der dannes et carbonyl af acetylderivatet cystein: cystein frigives, og der dannes et β-keto (acetylacetyl) phosphopantethinderivat.
Efterfølgende griber β-keto-acyl-ACP-reduktasen ind, hvilket reducerer carbonylet yderligere til ACP-enzymet (et hydroxid dannes af NADPH, som reduceres til NADP +).
Nu virker 3-hydroxy-acyl ACP dehydratase (dehydrering forekommer), hvilket fører til dannelsen af et umættet system (alken).
Den næste proces involverer enoyl-ACP-reduktase (den udfører en hydrogenering: alkanen dannes, og NADPH reduceres til NADP +).
Den sidste fase involverer omdannelse af acylproduktet opnået fra den første cyklus til en forbindelse, der er i stand til at starte en anden cyklus: transacylaseenzymet overfører acylen til cystein, hvilket efterlader stedet for pantethinen, som nu vil være villig til at binde en anden malonyl.
Ved β-oxidation anvendes et molekyle af FAD til at opnå det umættede α-β metabolit trans enoyl coenzym A ved dehydrogenering; ved syntese af fedtsyrer bruges der i stedet et molekyle af NADPH til at få den modsatte reaktion til at finde sted.
Normalt syntetiseres fedtsyrer med seksten carbonatomer, men fedtsyrer med atten, tyve eller toogtyve kulstofatomer kan også produceres; fedtsyrerne forestres derefter for at danne triglycerider med aktiveret glycerol (dvs. glycerol 3-phosphat). Sidstnævnte kan opnås fra dihydroxyacetonphosphat ved enzymets virkning glycerolphosphat dehydrogenase eller fra glycerol via enzymet glycerolkinase.
De syntetiserede fedtsyrer skal sendes til fedtvævet; de transporteres i blodbanen i form af triglycerider eller delvis som sådan ved brug af et transportprotein, som er albumin.