FORENING (ELLER "KONTAKT" ELLER "LINKAGE")
Indtil nu har vi behandlet den mendelske dihybrid (eller polyhybrid) krydsning under forudsætning af at de forskellige allelpar faktisk findes på forskellige homologe kromosompar. Men antallet af kromosomale par, selv om det er forskelligt fra art til art, varierer inden for snævre grænser (få arter når omkring hundrede kromosomer), mens antallet af gener kan tælles i titusinder.
At de karakterer, Mendel valgte til sine eksperimenter, adskilte alle uafhængigt (uden derfor at forvirre beregningerne i fordelingen af fænotyper i F2 i polyhybridkorset) var en heldig chance. Hvis der blev fundet to par alleler på tilstødende loci, var loven, der ville følge ville blive kaldt foreningsloven.
Når man ved, at rigtig mange tegn har deres locus på et enkelt kromosom, og at de er de kromosompar, der adskiller sig uafhængigt i meiose, kan det godt forstås, hvor ofte det sker, at to par tegn, hvis de var forbundet på et kromosom i forældrenes organisme, forbliver forbundet. ligeledes også i kønscellen og derfor i den organisme, som den vil bringe sit eget genetiske materiale til.
Således ser vi, at "forening repræsenterer en" undtagelse, langt fra sjælden, til den uafhængighed, der er beskrevet i Mendels tredje lov.
Bytte eller "krydse-over" og rekombination
Når vi taler om meiose, har vi angivet, at der er to forskellige øjeblikke af blanding af det genetiske materiale: det ene er adskillelsen af kromosomer i kønsceller, og det er det, der observeres af Mendel.
Det andet øjeblik, som faktisk går forud, er det, hvor de fire kromatider i hvert par af homologe kromosomer indbyrdes udveksler identiske træk. Efter denne udveksling vil to faktorer, der var forbundet på det samme kromosom i stedet være uafhængige i kønscellerne. Sandsynligheden at en udveksling finder sted er proportional med en første tilnærmelse til kromosomets længde, og i længere kromosomer kan der være endnu mere end en udveksling.
Fænomenet kan detekteres cytologisk ved at observere et tilstrækkeligt antal meioser under mikroskopet.
Genkampningshastigheden er den hastighed, hvormed to tegn, der blev associeret i forældregenerationen, rekombinerer forskelligt i F2.
Hvis de to loci er absolut sammenhængende, er sandsynligheden for, at en chiasme vil adskille dem, praktisk talt nul. Rekombinationshastigheden vil være: n ° rekombinanter. Hvis to loci er på to forskellige kromosomer, vil rekombinationshastigheden være 0,5 (lige sandsynlighed for to tegn, der blev forbundet i P -generationen, for tilfældigt at finde sig sammen i F2). Rekombinationshastigheden kan derfor variere mellem 0,0 og 0,5. For små afstande på kromosomet er afstanden og rekombinationshastigheden direkte proportional. Ved længere afstande er der mulighed for, at der finder to udvekslinger sted mellem to loci. Det vil nu fremstå klart, at to faktorer adskilt af to udvekslinger igen er forbundet. det er klart på dette tidspunkt, at proportionaliteten mellem lokalitetens afstand og sandsynligheden for rekombination er tabt.
Loci fundet forbundet på det samme kromosom udgør 'foreningsgrupper'. Meget fjerne loci kan have en sådan sandsynlighed for adskillelse ved udveksling, at de opfører sig som uafhængige, men hver af dem vil blive forbundet med en lavere rekombinationshastighed til de mellemliggende loci.
Når rekombinationshastighederne mellem mange par af gener inden for en foreningsgruppe kendes, kan konstruktionen af 'genetiske kort' begynde. Husk på, at afstanden mellem to gener (a og b) udtrykkes ved rekombinationshastigheden, og at afstanden fra a fra et tredje gen c enten kan være summen eller forskellen i forhold til dens afstand fra b, er det muligt at rekonstruere et kort over de gensidige afstande, som vil være det genetiske kort inden for den sammenslutningsgruppe, det vil sige det kromosom.
Vi skal nu generelt overveje nogle begreber, der begrænser den fænotypiske manifestation af genotypiske tegn.
Først og fremmest vil vi tale om begreberne penetration og udtryksfuldhed, og derefter vil vi lægge særlig vægt på fænomenerne regulering af genhandling.
PENETRANCE
Genets penetration repræsenterer dets evne til at manifestere sig i fænotypen. Penetrance måles statistisk ved at tælle hyppigheden af fænotyperne, der viser den karakter ud af 100 genotyper, der indeholder den. Et træk med 0,7 penetrans er et træk, der forekommer fænotypisk i 70% af dets genotypiske frekvens.
EXPRESSIVITET
Ekspressivitet er en kvantitativ vurdering af graden af fænotypisk manifestation.
FORORDNING OM GENEHANDLING
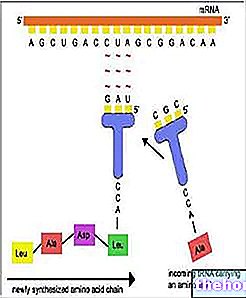
Celler producerer alle deres enzymer og proteiner med samme hastighed og på samme tid. Escherichia coli-celler kan f.eks. Forsynes med energi og carbonatomer fra lactosedisaccharidet, da de er i stand til at nedbryde dem til glucose og galactose takket være beta-galactosidase-enzymet. I et normalt E. coli, der kan have lactose, der er cirka 3000 molekyler beta-galactosidase, svarende til 3% af proteinerne i denne celle; i fravær af lactose vil der kun være ét molekyle beta-galactosidase pr. bakteriecelle. Galactosidase vil blive syntetiseret fra nye mRNA -molekyler, når den kan bruges. Mutantstammer af E. coli, der er rige på enzymet, kendes, selv når lactose mangler: Disse mutanter er dårligere stillet i forhold til normale celler, da de er tvunget til et unødigt forbrug af energi og materialer for at producere enzymet, der forbliver uden substrat. Stoffer, der forårsager en stigning i mængden af enzym, som det er tilfældet med lactose, vil blive kaldt inducere, mens enzymer siges at være inducerbare. Andre stoffer fremkalder, også disse på en bestemt måde, produktionen af visse enzymer. Også i E. coli, for eksempel i stand til at opbygge alle dets aminosyrer, der har carbon og ammonium (NH3), blokerer tilstedeværelsen i dyrkningsmediet af en bestemt aminosyre (f.eks. Histidin) produktionen af alle enzymer forbundet med selve aminosyrens biosyntese: det vil blive sagt om disse enzymer, at de er repressible. I bakterieceller nedbrydes mRNA -molekylerne kort efter deres dannelse, og det er derfor at kontrollere produktionen af mRNA betyder at kontrollere den enzymatiske syntese samtidig tid.
OPERONEN
For at forklare, hvordan bakteriecellen er i stand til at kontrollere sin egen produktion af enzymer, formulerede Jacob og Monod hypotesen om operonen; operonen dannes af flere gener, der er funktionelt relaterede og justeret uden diskontinuitet langs en strækning af DNA. Operonet består af tre forskellige typer gener: promotoren, hvor dannelsen af mRNA begynder; operatøren, hvor kontrollen udøves; et eller flere strukturelle gener, som koder for enzymer eller for andre proteiner. I beta-galactosidasesystemet omfatter operonet udover det for beta-galactosidase, også to andre gener strukturel kodning for andre enzymer involveret i metabolismen af lactose. Disse gener støder op til hinanden og transkriberes efter hinanden langs den samme DNA -helix til et enkelt mRNA -molekyle. De således producerede mRNA -molekyler er aktive i meget kort tid, hvorefter de ødelægges af specifikke enzymer.
Operonens aktivitet styres igen af et andet gen, regulatoren, som også kan være fjernt fra operonen: denne regulator koder for et protein, kaldet en repressor, som ser ud til at binde sig til DNA'et ved operatørgenet. mellem promotoren og de strukturelle gener blokerer faktisk produktionen af mRNA.
Repressoren styres igen, og kontrollen udføres ved hjælp af et "signal" -stof. I tilfælde af inducerbare enzymer er dette stof "induceren. Induceren" binder sig til repressormolekylet, der ændrer dets form, så det ikke længere kan tilpasse sig DNA'et: i dette tilfælde, da der ikke er nogen repressor mellem promotoren og strukturelle gener , kan repressoren danne mRNA -molekylerne og heraf proteinmolekylerne. Med udtømning af tilførslen af inducer igen vil regulatoren genvinde kontrollen, hvilket stopper produktionen af nyt mRNA, derfor af nye proteiner.I beta-galactosidasesystemet er induceren lactose eller et stof, der ligner meget dette. Derivat: de vil slutte sig til repressoren og inaktivere den for at muliggøre biosyntese af enzymer. I tilfælde af repressible enzymer fungerer stoffet, der fungerer som et "signal" som en corepressor: repressoren er kun aktiv, hvis den kombineres med corepressoren. I histidinsystemet, der involverer et dusin forskellige enzymer, er det denne aminosyre kombineret med dens tRNA, corepressoren. Histidin.
ALLOSTERISKE INTERAKTIONER
Allosteriske interaktioner, der involverer inaktivering af et enzym ved at ændre dets form, giver en anden måde at regulere en celles metaboliske aktivitet på. Allosteriske interaktioner giver mulighed for mere præcis kontrol end operonets induktor -repressorsystem, men opnår ikke det nyttige resultat ved at udelukke biosyntesen af et givet stof fra det første trin - produktionen af et mRNA.
KONTROLSYSTEMER I EUCARIUS
Der er nogle kendsgerninger, der får en til at tro, at et reguleringssystem, der ligner operonen, fungerer og er fremtrædende blandt planter og dyr.Disse organismeres kromosomer adskiller sig meget fra E. coli og andre prokaryoter. Kontrollen med generne i disse celler er meget forskellige. Mitosemekanismen er sådan, at hver celle i en given plante eller et dyr besidder al information
genetik til stede i det befrugtede æg. Derfor vil de fleste gener i enhver specialiseret celle forblive ineffektiv i hele cellens levetid. DNA'et i disse celler er altid forbundet med proteiner. Så det er muligt, at genrepression i eukaryoter kræver netop denne sammenhæng mellem DNA og proteiner.