Redigeret af Dr. Giovanni Chetta
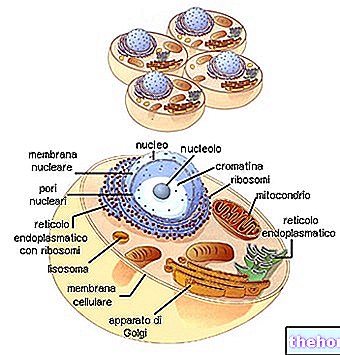
ECM beskrives generelt som værende sammensat af flere store klasser af biomolekyler:
- Strukturelle proteiner (kollagen og elastin)
- Specialiserede proteiner (fibrillin, fibronectin, laminin osv.)
- Proteoglycaner (aggrecaner, syndecaner) og glusaminoglycaner (hyaluronaner, chondroitinsulfater, heparansulfater osv.)
Strukturelle proteiner
Kollagener danner den mest repræsenterede familie af glykoproteiner i dyreriget. De er de mest tilstedeværende proteiner i den ekstracellulære matrix (men ikke den vigtigste) og er de grundlæggende bestanddele i det rigtige bindevæv (brusk, knogle, fascia, sener, ledbånd).
Der er mindst 16 forskellige typer kollagen, hvoraf type I, II og III er de mest tilstedeværende på niveau med de typiske fibriller (type IV danner en slags retikulum, der repræsenterer hovedkomponenten i de basale laminer).
Kollagener er for det meste syntetiseret af fibroblaster, men epitelceller er også i stand til at syntetisere dem.
Kollagenfibre interagerer kontinuerligt med en enorm mængde andre molekyler i den ekstracellulære matrix, hvilket udgør et biologisk kontinuum, der er grundlæggende for cellens liv.De associerede kollagener i fibriller spiller en dominerende rolle i dannelsen og vedligeholdelsen af strukturer, der er i stand til at modstå spændingskræfter, er næsten uelastisk (glucosaminglycaner udfører en handling af modstandsdygtighed over for kompression). På en eller anden måde produceres og metaboliseres kollagen igen som en funktion af den mekaniske belastning og dens viskoelastiske egenskaber, som vi vil se i afsnittet "Viskoelasticitet af fascia ", en stor indvirkning på menneskets kropsholdning. Som en yderligere demonstration af kollagens evne til at ændre sig efter miljøpåvirkninger, forudsat f.eks. varierende grader af stivhed, elasticitet og modstand, er der kollagener, defineret med udtrykket FACIT (Fibril Associated Collagen with Interrupted Triple helices) i stand til at fungere funktionelt som proteoglycaner (beskrevet i afsnittet "Glucosaminoglycaner og proteoglycaner").
Kollagenfibrene har takket være deres belægning af PG / GAG (proteoglycaner / glucosaminoglycaner) egenskaber ved biosensorer og bioledere: de relative elektriske ladninger resulterer i en større evne til at binde vand og udveksle ioner, derfor en større elektrisk kapacitet.
Vi ved, at enhver mekanisk kraft, der er i stand til at generere en strukturel deformation, stresser de intermolekylære bindinger og producerer en lille elektrisk flux, dvs. piezoelektrisk strøm (Athenstaedt, 1969). I sådanne tilfælde fordeler kollagenfibrene de positive ladninger på deres konvekse overflade og de negative på den konkave og transformerer således til halvledere (de tillader strømning af elektroner på deres envejs overflade). Da den piezoelektriske energi (såvel som den pyroelektriske energi genereret af termiske spændinger) neutraliseres af de cirkulerende ioner på meget kort tid (ca. 10-7-10-9 sekunder), er arrangementet af PG / GAG på signalet er afgørende for spredning af signaloverfladen på fibrillerne, for eksempel at virke som "repeatere" af den elektriske impuls. Især en langsgående periodicitet på ca. 64 nm (som under det optiske mikroskop optræder som en streak) tillader en forplantningshastighed af impulsen svarende til ca. 64 m / s (svarende til ledningshastigheden for hurtige nervefibre) - Rengling, 2001. Det stærke dipolære moment af kollagenfibriller og deres resonanskapacitet (egenskab, der er fælles for alle peptidstrukturer), såvel som MEC's lave dielektriske konstant, letter transmissionen af elektromagnetiske signaler.Derfor besidder det tredimensionale og allestedsnærværende kollagennet også den særegne egenskab ved at lede bioelektriske signaler i de tre dimensioner af rummet, baseret på det relative arrangement mellem kollagenfibriller og celler, i afferent retning (fra ECM til cellerne) eller omvendt efferent.
Alt dette repræsenterer et real-time MEC-celle kommunikationssystem, og sådanne elektromagnetiske bio-signaler kan føre til vigtige biokemiske ændringer, for eksempel i knogler kan osteoklaster ikke "fordøje" piezoelektrisk ladet knogle (Oschman, 2000).
Endelig skal det understreges, at cellen, ikke overraskende, producerer kontinuerligt og med et betydeligt energiforbrug (ca. 70%) materiale, der nødvendigvis skal udvises, hovedsageligt gennem den eksklusive opbevaring af protocollagen (biologisk forløber for kollagen) i specifikke vesikler (Albergati, 2004).
Langt de fleste hvirveldyr væv kræver samtidig tilstedeværelse af to vitale egenskaber: styrke og elasticitet. Et reelt netværk af elastiske fibre, der er placeret inde i ECM for disse væv, gør det muligt at vende tilbage til de oprindelige forhold efter stærke trækninger.De elastiske fibre er i stand til at øge udstrækningen af et organ eller en del af det med mindst fem gange. Lange, uelastiske kollagenfibre er spredt mellem de elastiske fibre med den præcise opgave at begrænse "overdreven deformation på grund af træk i vævene. L"elastin repræsenterer hovedkomponenten i elastiske fibre. Det er et ekstremt hydrofobt protein, cirka 750 aminosyrer i længden, da kollagen er rig på prolin og glycin, men i modsætning til kollagen er det ikke glyceret og indeholder mange hydroxyprolinrester og ikke hydroxylisin. Elastin fremstår som et rigtigt biokemisk netværk af uregelmæssigt tredimensionel form, sammensat af fibre og lameller, der gennemsyrer ECM for alle bindevæv. Det findes i særligt rigelige mængder i blodkarrene med elastiske egenskaber (det er proteinet af ECM mere til stede i arterierne og repræsenterer mere end 50% af aortas samlede tørvægt), i ledbåndene, i lungen og i huden. I dermis, i modsætning til hvad der sker med kollagen, har densitet og volumen af elastin en tendens til at stige over tid, men det gamle elastin fremstår generelt hævet, næsten hævet, ofte med et fragmenteret udseende og med en reduktion i komponenten. "Amorf" (Pasquali Rochetti et al, 2004). Glatte muskelceller og fibroblaster er de største producenter af dens forløber, tropoelastin, udskilt i de ekstracellulære rum.
Andre artikler om "Kollagen og elastin, kollagenfibre i den ekstracellulære matrix"
- Ekstracellulær matrix
- Fibronectin, Glucosaminoglycaner og Proteoglycaner
- Betydningen af den ekstracellulære matrix i cellulære ligevægte
- Ændringer af den ekstracellulære matrix og patologier
- Bindevæv og ekstracellulær matrix
- Deep fascia - Bindevæv
- Fasciale mekanoreceptorer og myofibroblaster
- Deep fascia biomekanik
- Holdning og dynamisk balance
- Tensegrity og spiralformede bevægelser
- Underekstremiteter og kropsbevægelse
- Støttestøtte og stomatognatisk apparat
- Kliniske tilfælde, postural ændringer
- Kliniske tilfælde, kropsholdning
- Postural evaluering - Klinisk case
- Bibliografi - Fra den ekstracellulære matrix til kropsholdning. Er forbindelsessystemet vores sande Deus ex machina?